General
- S6: Biology SB File Uploaded 3/08/22, 12:46
- S6: Biology TG File Uploaded 3/08/22, 12:49
UNIT 13: PRINCIPLES OF GENE TECHNOLOGY
Key Unit Competence
Explain the principles of gene technology.Learning Objectives
By the end of this unit, I should be able to:
–– Define the term recombinant DNA.
–– Explain that genetic engineering involves the extraction of genes from one organism or the synthesis of genes, in order to place them in another organism (of the same or another species) such that the receiving organism expresses the gene product.
–– Describe the properties of plasmids that allow them to be used in gene cloning.
–– Explain the use of genes in fluorescent or easily stained substances as markers in gene technology.
–– Describe the principles of the Polymerase Chain Reaction (PCR) to clone and amplify DNA (the role of Taq polymerase should be emphasized).
–– Describe and explain how gel electrophoresis is used to analyse proteins and nucleic acids, and to distinguish between the alleles of a gene (limited to the separation of polypeptides and the separation of DNA fragments cut with restriction endonucleases).
–– Explain the roles of restriction endonucleases, reverse transcriptase and ligases in genetic engineering.
–– Explain and outline, how microarrays are used in the analysis of genomes and in detecting mRNA in studies of gene expression.
–– Interpret illustrations of the isolation and transfer of genes using plasmids in transgenic organisms (bacteria, plant or an animal).
–– Sequence the processes involved in the extraction and transfer of genes from one organism to another.
–– Interpret charts of the Polymerase Chain Reaction (PCR).
–– Relate the mechanism of DNA replication to PCR and the amount of DNA produced in a given period of time.
–– Appreciate that the easy transfer of some plastids from one species of bacteria to another may carry genes for antibiotic resistance.
–– Acknowledge that advances in genetic engineering have enabled manipulation of genes to our advantage
13.1 Recombinant DNA and enzymes involved in genetic engineering

13.1.1 Recombinant DNA
A recombinant deoxyribonucleic acid (r DNA) is the DNA that contains genes from more than one source. Examples of molecules produced from recombinant DNA and that are important to humans include some pharmaceuticals like human insulin and antibiotics.
Genetic engineering, also known as recombinant DNA technology or gene cloning or gene technology is the alteration of the genes in a living organism to produce a genetically modified organism (GMO) with a new genotype. Various kinds of genetic modification are possible and include:
–– Inserting a foreign gene from one species into another in order to form a transgenic organism,
–– Altering an existing gene so that its product is changed and changing gene expression so that it is translated more often or not at all.13.1.2 Role of some enzymes in genetic engineering
The enzymes involved in gene manipulation include; restriction endonucleases (restriction enzymes), methylase, ligase and reverse transcriptase.
a. Restriction endonucleases
Different restriction enzymes, also called restriction endonucleases, exist and cut the DNA molecule into fragments; their examples are shown in the table 13.1below.
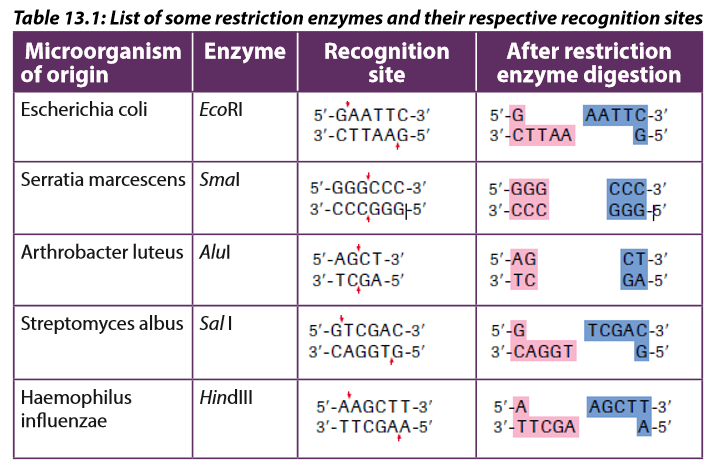
Restriction enzymes are named according to the bacteria from which they originate.
For example, the restriction enzyme BamHI is named as follows:
–– B represents the genus Bacillus
–– am represents the species amyloliquefaciens–– H represents the strain
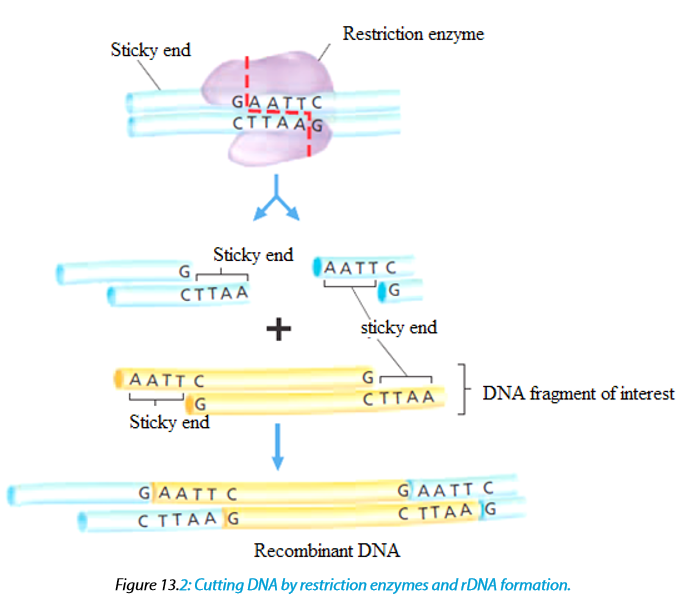
–– I mean that it was the first endonuclease isolated from this strainA commonly used tool in molecular biology is restriction endonucleases which are molecular scissors that can cut double-stranded DNA at a specific base-pair sequence. Each type of restriction enzyme recognizes a characteristic sequence of nucleotides that is known as its recognition site. Most recognition sites are four to eight base pairs long and are usually characterized by a complementary palindromic sequence.
For example, the restriction enzyme EcoRI binds to the following base-pair sequence: 5’-GAATTC-3’/3’-CTTAAG-5’. It is palindromic because both strands have the same base sequence when read in the 5’ to 3’ direction. EcoRI scans a DNA molecule and only stops when it is able to bind to its recognition site. Once bound, it disrupts, via a hydrolysis reaction, the phosphodiester bond between the guanine and adenine nucleotides on each strand. A phosphodiester bond is a covalent bond located between a two sugar groups and a phosphate group; such bonds form the sugar-phosphate backbone of DNA and RNA. Subsequently, the hydrogen bonds of complementary base pairs between the cuts are disrupted. The result is a cut within a DNA strand, producing two DNA fragments where once there was only one.
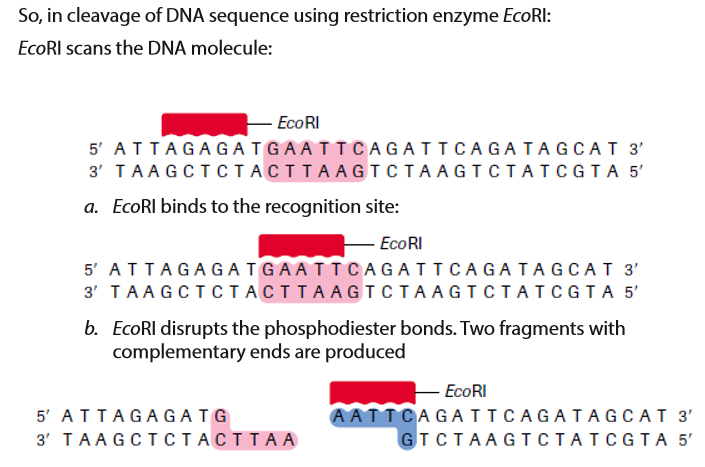
The ends of DNA fragments produced from a cut by different restriction endonucleases differ, depending on where the phosphodiester bonds are broken in the recognition site. In the example in Table 13.1, EcoRI produces sticky ends; that is, both fragments have DNA nucleotides that are now lacking their respective complementary bases. These overhangs are produced because EcoRI cleaves between the guanine and the adenine nucleotide on each strand. Since A and G are at opposite ends of the recognition site on each of the complementary strands, the result is the overhang. In few words, sticky ends are fragment end of a DNA molecule with short single stranded overhangs, resulting from cleavage by a restriction enzyme
b. Methylases
These are enzymes that add a methyl group (CH3) to one of the nucleotides found in a restriction endonuclease recognition site, altering its chemical composition. They allow the molecular biologist to protect a gene fragment from being cleaved in an undesired location.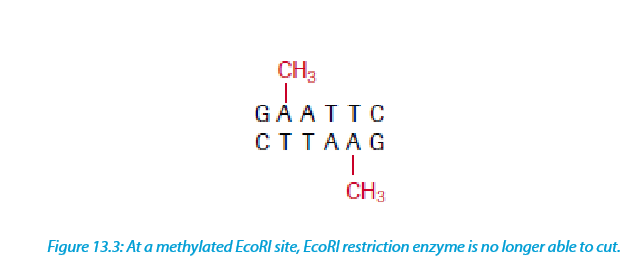
c. DNA ligase
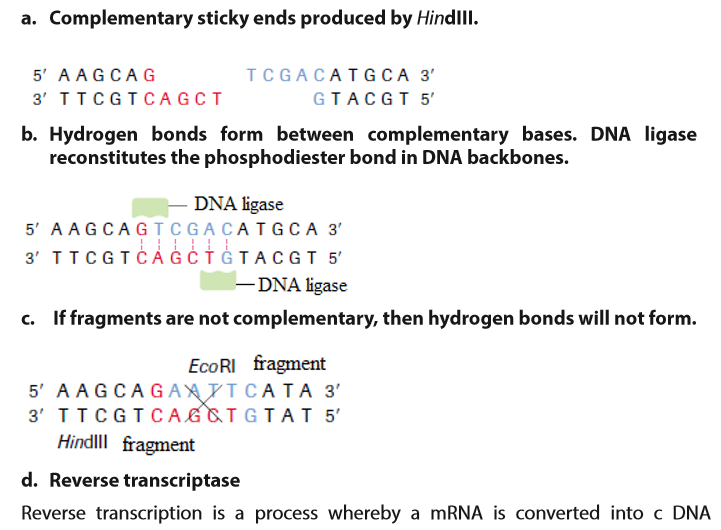
This enzyme repairs broken DNA by joining two nucleotides in a DNA strand. It is commonly used in genetic engineering to do the reverse of a restriction enzyme that is to join together complementary restriction fragments. The sticky ends allow two complementary restriction fragments to harden, but only by weak hydrogen bonds, which can quite easily be broken by gentle heating. The backbone is still incomplete. DNA ligase completes the DNA backbone by forming covalent bonds. T4 DNA ligase is an enzyme that originated from the T4 bacteriophage and which is used to join together DNA blunt or sticky ends. So, DNA ligase is able to join complementary sticky ends produced by the same restriction enzyme via a condensation reaction:

13.2 Properties of plasmids and gene manipulation

13.2.1. Properties of plasmids
A plasmid is a genetic structure, in some cells, that can replicate independently of the chromosomes; it is typically a small circular DNA strand in the cytoplasm of a bacterium or protozoan. Plasmids are much used in the laboratory during manipulation of genes.

The properties of plasmids are:
–– It is big enough to hold the desired gene.
–– It is circular (or more accurately a closed loop), so that it is less likely to be broken down.
–– It contains control sequences, such as a transcription promoter, so that the gene will be replicated or expressed.
–– It contains marker genes, so that cells containing the vector can be identified.Plasmids are not the only type of vector that can be used. Viruses can also be used as vectors. Another group of vectors are liposomes, which are tiny spheres of lipid containing the DNA.
13.2.2. Gene manipulation
Genetic manipulation is a process done to use the genome of an organism in order to produce desired traits. A genome is the complete set of genes or genetic material present in an organism.
Genes are pieces of DNA, that carry carrier of the genetic information which determines all the characteristics of an individual such as eye colour, size, ability to resist disease, etc. Gene manipulation involves mainly the transfer of genes from one organism to another.
The overview of gene transfer, resulting in genetically modified organisms (GMO) also called transgenic organisms such as bacteria or animals or plants having foreign gene inserted into them, is shown below:
1. Generation of DNA fragments using restriction endonucleases:
–– Appropriate restriction endonucleases need to be used to ensure that the gene fragment in question is excised completely from the source DNA.–– More than one restriction endonuclease may be used at one time.
2. Construction of a recombinant DNA molecule:
–– The target gene fragment is ligated to a DNA vector (plasmids are one example) and is now recombinant DNA.
–– The vector can replicate autonomously in an appropriate host organism.
3. Introduction into a host cell:
–– Bacterial host cells can be manipulated to take up the recombinant DNA using electroporators, gene guns or classical transformation protocols.
–– Once the bacterium takes up the recombinant DNA, it is referred to as being transformed.
4. Selection:
–– Cells that have been successfully transformed with the recombinant DNA must be isolated.
–– The desired cells are usually chemically selected by the presence of a marker (e.g. antibiotic resistance) on the vector.
–– Growth of colonies on media containing the chemical indicates successful transformation of the recombinant DNA vector.
–– Individual colonies are isolated from media containing the chemical and are grown in culture to produce multiple copies (clones) of the incorporated recombinant DNA. Different gene manipulations are illustrated under the heading 13.3.
To perform these gene manipulation steps, the genetic engineer needs a tool kit consisting of:
1. Enzymes, such as restriction endonucleases (restriction enzymes), ligase and reverse transcriptase
2. Vectors, including plasmids and viruses
3. Genes coding for easily identifiable substances that can be used as markers.
13.3 Transfer of genes using plasmids in transgenic organisms

The production of genetically modified organisms (GMO), also called transgenic
organisms, is a multistage process which can be generally summarized as follows:
–– Identification of the gene of interest.
–– Isolation of the gene of interest.
–– Cutting of gene of interest and opening of plasmid with restriction enzymes in
order to have sticky ends
–– Associating the gene with an appropriate promoter and poly -A sequence and
insertion into plasmids.
–– Multiplying the plasmid in bacteria and recovering the cloned construct for injection.
–– Transference of the construct into the recipient tissue, usually fertilized eggs.
–– Integration of gene into recipient genome.
–– Expression of gene in recipient genome.
–– Inheritance of gene through further generations.13.3.1. Extraction, purification, isolation and transfer of genes using plasmids into bacteria
The normal gene coding for a particular protein is extracted from an organism; it is isolated and transferred into a plasmid of a bacterium. This plasmid becomes a recombinant DNA that is introduced into that bacterium. This bacterium becomes a transgenic bacterium. An example of the sequence of the processes involved in the extraction and transfer of genes from one organism to another is illustrated below.
Process 1: Extraction and purification of DNA containing an interest gene is required for a variety of molecular biology applications. Its process is summarized below.

The purification of DNA from cell extract occurs in this way:
–– The standard way to deproteinize a cell is to add phenol or a 1:1 mixture of phenol and chloroform.
–– The organic solvents precipitate proteins but leave the nucleic acids (DNA and RNA) in an aqueous solution.
–– The result is that is the cell extract is mixed gently with the solvent, and the layers then separated by centrifugation, precipitated protein molecules are left as a white coagulated mass at the interface between the aqueous and organic layers.
–– The aqueous solution of nucleic acids can then be removed with a white pipette.
–– Cell extract is treated with protease such as pronase or proteinase K before extraction.
–– These enzymes will break polypeptides into smaller units thus making phenol easier to remove them.
–– The only effective way to get rid of RNA is the use of ribonuclease enzyme which will rapidly degrade the molecules into ribonucleotide subunits. As DNA is purified, also its genes are purified.
The Concentration of DNA samples is carried out in this way:
–– The most frequently used method of concentration is ethanol precipitation.
–– In the presence of salt and a temperature of -20 oC or less absolute ethanol
with efficiently precipitate polymeric nucleic acids.
–– With 2 thick solution of DNA, the ethanol can be layered on the top of the sample.
–– A spectacular trick is to push a glass rod through the ethanol into the DNA solution.
–– When the rod is removed, DNA molecules will adhere and be pulled out of the solution in the form of long fiber.
After getting DNA, it is possible to remove the gene from it, by a restriction enzyme, in order to use it for a particular purpose. For example, normal insulin gene is removed from human cell as shown in the figure below.

Process 2: Summary of transfer of insulin gene using plasmids into bacteria

The plasmid is now an example of recombinant DNA, which can be introduced into a bacterial cell to produce numerous copies (clones) of the gene. As the inserted gene codes for insulin, a hormone that reduces the blood glucose level, and this gene functions normally as expected, the product (insulin) may also be retrieved and used for therapeutic purposes in which it is given to diabetic people.
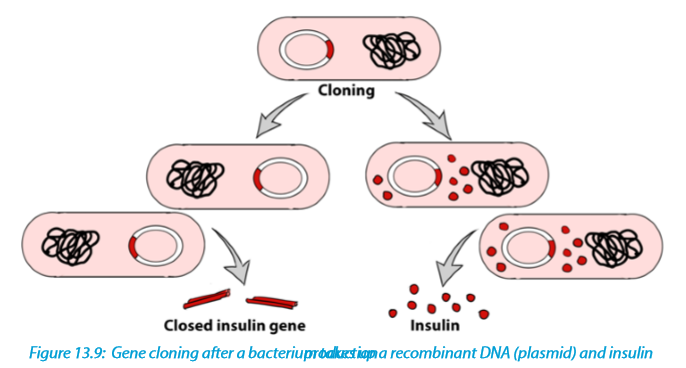
In nuclear biology and molecular biology, a marker gene is a gene used to determine if a nucleic acid sequence has been successfully inserted into an organism›s DNA.
13.3.2 Use of Agrobacterium tumefaciens to transfer genes in plants
Agrobacterium is a bacterium that uses a horizontal gene transfer (HGT). HGT is the transfer of DNA between different genomes. HGT can occur in bacteria through transformation, conjugation and transduction. However, it is also possible for HGT to occur between eukaryotes and bacteria. Bacteria have three ways of transferring bacteria DNA between cells:
1. Transformation: The uptake and incorporation of external DNA into the cell thereby resulting in the alteration of the genome.
2. Conjugation: The exchange of genetic material through cell-to-cell contact of two bacterial cells. A strand of plasmid DNA is transferred to the recipient cell and the donor cell then synthesis DNA to replace the strand that was transferred to the recipient cell.
3. Transduction: A segment of bacterial DNA is carried from one bacterial cell to another by a bacteriophage. The bacteriophage infects a bacterial cell and takes up bacterial DNA. When this phage infects another cell, it transfers the bacterial DNA to the new cell. The bacteria can then become a part of the new host cell.Agrobacterium also has the ability to transfer DNA between itself and plants and is therefore commonly used in genetic engineering. The process of using Agrobacterium for genetic engineering is illustrated in the diagram below.
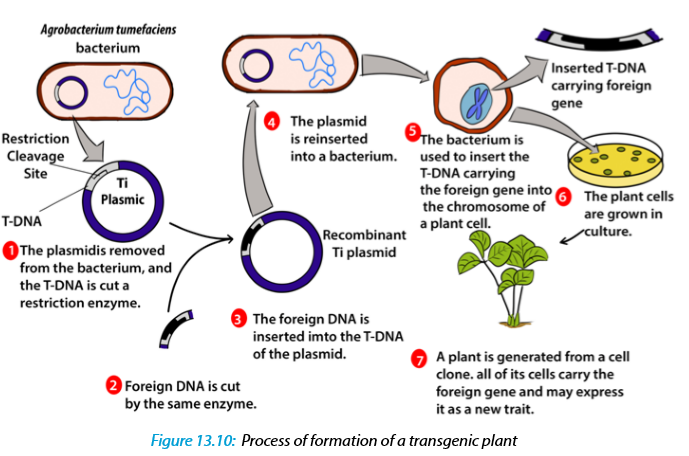
Summary of formation of a transgenic plant:
–– The agrobacterium cell contains a bacterial chromosome and a Tumor inducing plasmid (Ti Plasmid).
–– The Ti plasmid is removed from the agrobacterium cell and a restriction enzyme cleaves the T-DNA restriction site. The transfer DNA (T-DNA) is the transferred DNA of the tumor-inducing plasmid of some species of bacteria such as Agrobacterium tumefaciens
–– The T-DNA is transferred from bacterium into the host plant›s nuclear DNA genome.
–– Next foreign DNA, which is also cleaved by the same enzyme, is inserted into the T -DNA at the site that was cleavage site.
–– The modified plasmid is then reinserted in the agrobacterium and the bacterium inserts the T-DNA, which now carries a foreign gene into the plant cell.
–– The plant cell is then cultured and results in a new plant that has the foreign DNA trait.13.3.3 Transfer of genes into animals
In reproductive cloning, researchers remove a mature somatic cell, such as a skin cell, from an animal that they wish to copy. They then transfer the DNA of the donor animal’s somatic cell into an egg cell, or oocyte, that has had its own DNA-containing nucleus removed. For example, the cell used as the donor for the cloning of Dolly sheep was taken from a mammary gland and the production of a healthy clone therefore proved that a cell taken from a specific part of the body could recreate a whole individual.
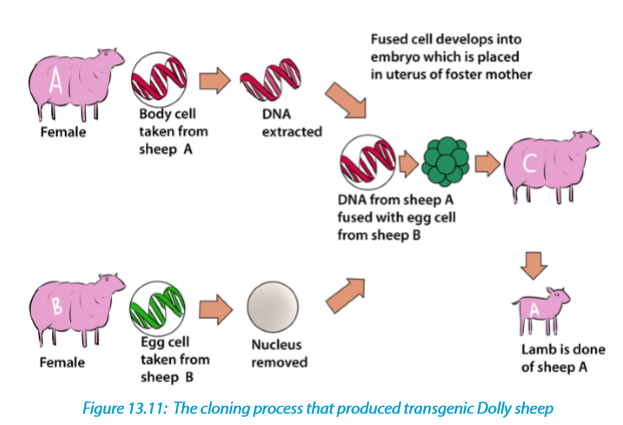
13.3.4 Transformation of harmless bacteria to pathogenic bacteria and resistant bacteria
A pathogenic bacterium is a bacterium which is capable of causing a disease. An example of harmful or pathogenic bacterium is Vibrio cholerae which causes cholera. A harmless bacterium can become pathogenic bacterium due to certain factors. The discovery of DNA and the genetic code led scientists to determine that some bacteria were resistant to particular antibiotics because of inserted genes that rendered bacteria unaffected by the effects of some antibiotics. This gene insertion can be done naturally between bacteria or artificially by biotechnologists.
Antibiotic resistance, also known as drug resistance, is the ability of bacteria and other microorganisms to resist the effects of an antibiotic to which they were once sensitive. Since bacteria are ubiquitous in the colon, conjugation is constantly occurring. This conclusion has been supported by bacteria in different genera containing homologous DNA plasmids. Therefore, horizontal gene transfer can occur between different species or within a population. This can become problematic if harmful bacteria that have been artificially selected for antibiotic resistance happen to be in the colon, where bacteria can transfer the resistance gene to other species of bacteria. Typically, this is not a problem because most bacteria are not harmful, unless bacteria that are a public health concern happen to receive a resistance gene.
Individuals that have previously taken antibiotics are less responsive to treatment because their bodies contain more antibiotic resistant bacteria.
These bacteria received these genes from disease - causing microbes that transferred a resistance gene through conjugation or transformation. The harmless bacteria that are resistant to antibiotics can then pass this gene to harmful bacteria that do not yet have antibiotic resistance. Thus, horizontal gene transfer allows bacteria to indirectly become resistant to antibiotics. Transformation and conjugation contribute to increasing frequencies of antibiotic resistant genes because of genes transferring between different species. The gene transfer can transform harmless bacteria into pathogenic bacteria which can cause diseases.
The prevention of antibiotic-resistant infections includes:
–– Do not take antibiotics for viral infections.
–– Complete your prescribed course of treatment exactly as instructed by your healthcare provider. Do not stop taking your medicine even if you feel better, and do not save any antibiotics for future use.
–– Do not take someone else’s antibiotics because different kinds of antibiotics treat different types of bacterial infections.
13.4 Non-biological methods of gene transfer

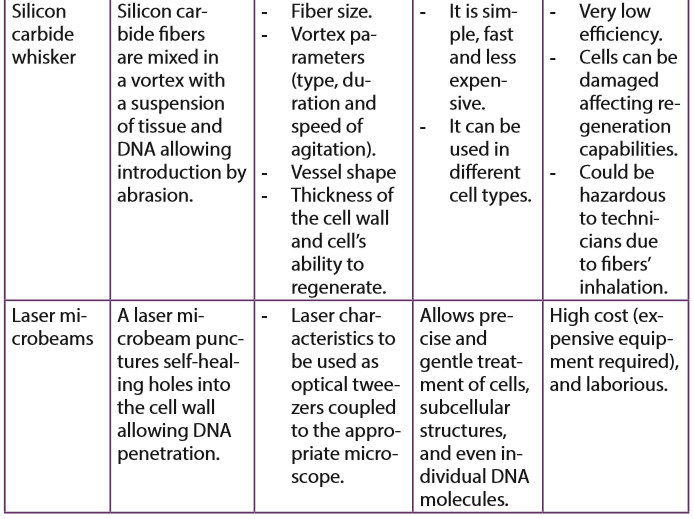
Different non-biological methods, also called physical methods or direct methods of gene transfer exist and include genetic transformation, shock wave-mediated genetic transformation, electroporation, biolistic, vacuum infiltration, silicon carbide whisker and laser microbeams.
a. Electroporation
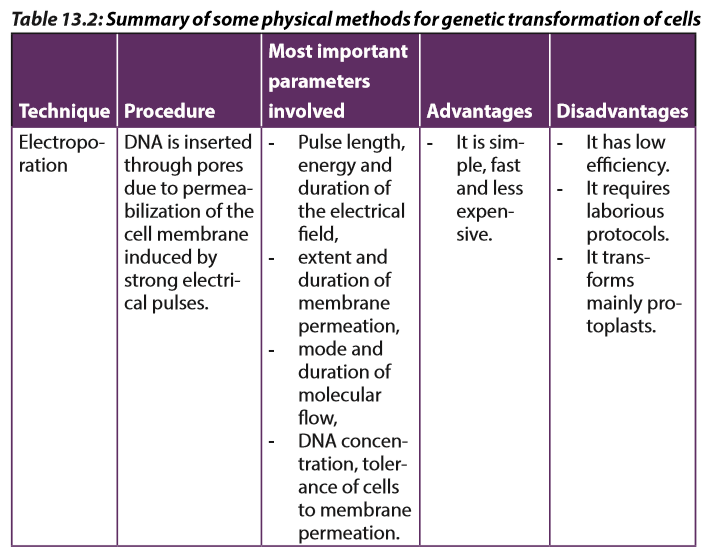
Electroporation is a method of transformation via direct gene transfer. In this technique, a mixture containing cells and DNA is exposed to very high voltage electrical pulses (4000 – 8000 Volts/cm) for very brief time periods (few milliseconds). It results in formation of transient pores in the plasma membrane, through which DNA seems to enter inside the cell and then nucleus.

A suspension of cells with plasmid DNA is taken in an electroporation cuvette placed between electrodes and electrical pulses are applied. Temporary micropores are formed in cell membranes which allow cells to take up plasmid DNA leading to stable or transient DNA expression.
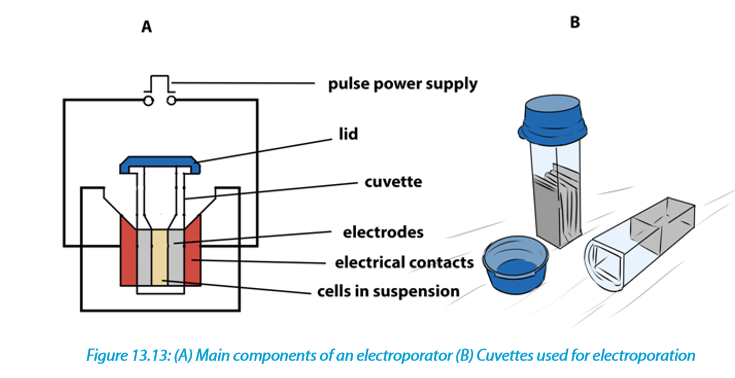
Cells which are arrested at metaphase stage of cell cycle are especially suitable for electroporation as these cells have absence of nuclear envelope and an unusual permeability of the plasma membrane. Protoplasts are used for electroporation of plant cells as thick plant cell walls restrict movement of DNA. The electroporation method was originally developed for protoplasts, but has given equally good results with cells and even tissues with easy recovery of regenerated plantlets. Immature zygotic embryos and embryogenic cells have also been used for electroporation to produce transgenic maize.
Transformation of protoplast is associated with low transient expression of transgenes as compared to organized tissues and low regeneration frequency especially in monocotyledonous plants. The electrical field and chemical substances applied to disorganize cell walls reduce the viability and capability of division of protoplasts.
b. Biolistics
Biolistics, also called micro projectile or particle bombardment, is a method where cells are physically impregnated with nucleic acids or other biological molecules.
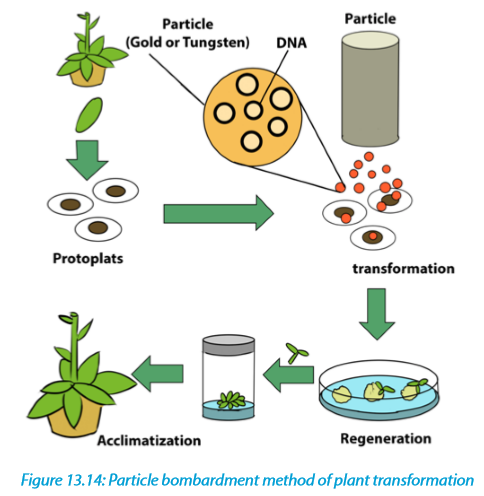
The main steps of a biolistic method are:
–– Isolation of protoplasts.
–– Injection of DNA-coated particles using particle gun.
–– Regeneration of transformed protoplasts into plantlets.
–– Acclimatization of regenerated plantlets in a greenhouse.A biolistic particle delivery system is a device for plant transformation where cells are bombarded with heavy metal particles coated with DNA/RNA. This technique was invented by John Stanford in 1984 for introduction of DNA into cells by physical means to avoid the host-range restrictions of Agrobacterium. Agrobacteriummediated genetic transformation system works well for dicotyledonous plants but has low efficiency for monocots. Biolistic particle delivery system provides an effective and versatile way to transform almost all type of cells. It has been proven to be a successful alternative for creating transgenic organisms in prokaryotes, mammalian and plant species.
c. Microinjection
The process of using a fine glass micropipette to manually inject transgene at microscopic or borderline macroscopic level is known as microinjection.
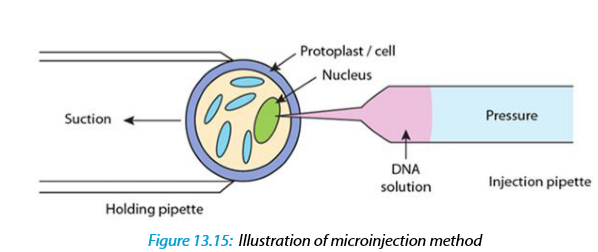
The transgene, in the form of plasmids, cosmids, phage or PCR products, can be circular or linear and does not need to be physically linked for injection. Microinjection involves direct mechanical introduction of DNA into the nucleus or cytoplasm using a glass microcapillary injection pipette. The protoplasts are immobilized in low melting agar, while working under a microscope, using a holding pipette and suction force. DNA is then directly injected into the cytoplasm or the nucleus. The injected cells are then cultured in vitro and regenerated into plants. Successful examples of this process have been shown in rapeseed, tobacco and various other plants.
Stable transformants can be achieved through this method but it requires technical expertise and is a time consuming process. Also, microinjection has achieved only limited success in plant transformation due to the thick cell walls of plants and a lack of availability of a single-cell-to-plant regeneration system in most plant species. In this technique, a traditional compound microscope (around 200x magnification) or an inverted microscope (around 200x magnification) or a dissecting stereomicroscope (around 40-50x) is used. The microscope target cell is positioned, cell membrane and nuclear envelope are penetrated with the help of two micromanipulators. One micromanipulator holds the pipette and another holds the micro capillary needle.
The two types of microinjection systems are constant flow system and pulsed flow system.
–– In the constant flow system, the amount of sample injected is determined by the duration for which needle remains in the cell. The constant flow system is relatively simple and inexpensive but outdated.–– The pulsed flow system has greater control over the volume of substance delivered, needle placement and movement and has better precision. This technique results in less damage to the receiving cell; however, the components of this system are quite expensive.
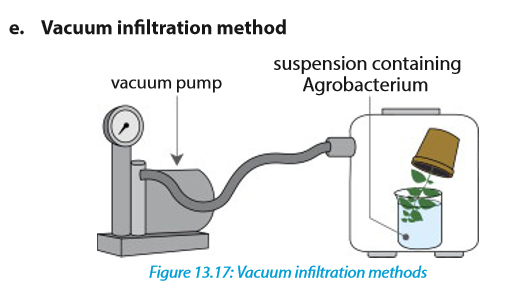
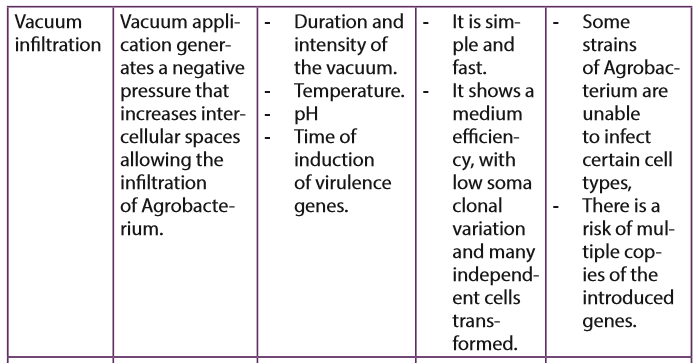
In this method, a vacuum pump generates a negative pressure that increases intercellular spaces allowing the infiltration of Agrobacterium.
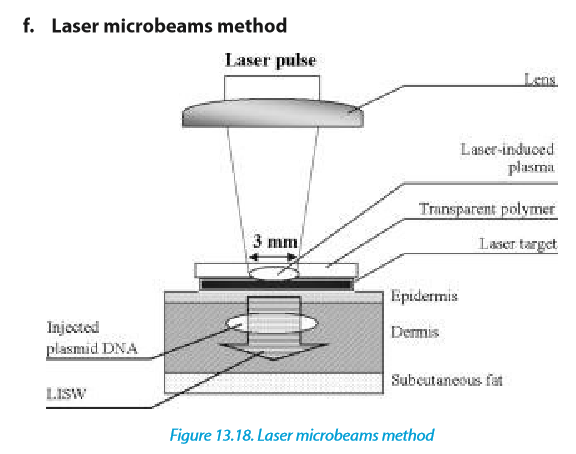
In this method, a laser microbeam punctures self-healing holes into the cell wall allowing DNA penetration.
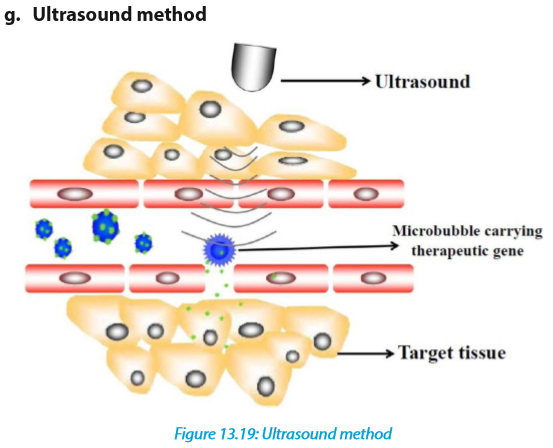
This method requires that a gene introduces DNA molecules into cells via acoustic cavitation that temporarily changes the permeability of the cell membrane. Sonoporation, or cellular sonication, is the use of sound (typically ultrasonic frequencies) for modifying the permeability of the cell plasma membrane. This technique is usually used in molecular biology and non-viral gene therapy in order to allow uptake of large molecules such as DNA into the cell, in a cell disruption process called transfection or transformation. Sonoporation employs the acoustic cavitation of microbubbles to enhance delivery of these large molecules. Sonoporation is under active study for the introduction of foreign genes in tissue culture cells, especially mammalian cells. Sonoporation is also being studied for use in targeted gene therapy in vivo, in a medical treatment scenario whereby a patient is given modified DNA, and an ultrasonic transducer might target this modified DNA into specific regions of the patient’s body.
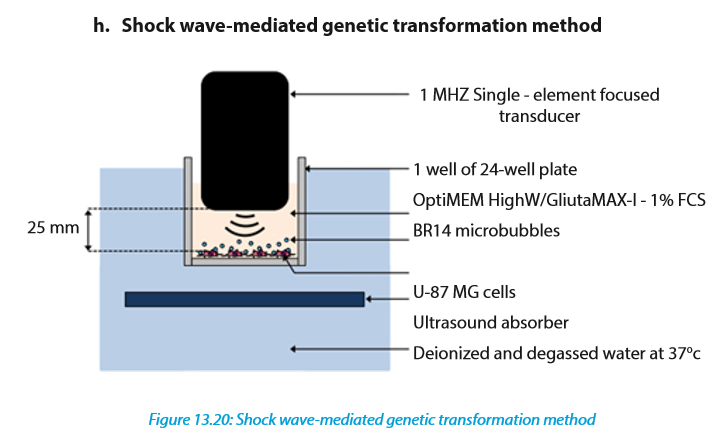
This method involves sonoporation, based on the use of high frequency ultrasound (1–10 MHz) in combination with gas microbubbles was introduced as a non-viral physical method that is currently under evaluation for gene and drug delivery. Sonoporation involves the treatment of a desired volume of cells in vitro or tissue in vivo with ultrasound in the presence of microbubbles. These microbubbles, which are formulated as lipid, albumin or polymer shelled micrometer sized gas bodies in aqueous suspension, are commonly mixed with cells for in vitro applications or administered by intravascular or intratissue injection for in vivo applications. The exposure of microbubbles to ultrasound causes their periodic oscillations and/or their collapse, under appropriate insonation conditions. It is now known that these oscillations can induce micro-streaming, shock waves and/or micro-jets that can affect the integrity of biological barriers (e.g. cell membrane, endothelial barrier). The use of sonoporation to deliver therapeutic molecules to tissues has been extensively explored over the past decade.
For example, the loco-regional delivery of anti-tumoral drugs has been reported and is now under clinical investigation. Sonoporation has been successfully used to transfer nucleic acids such as DNA into the heart, skeletal muscle, tumors, vessels, liver and kidney. This method enables exogenous delivery of molecules with minimal cell or tissue damage, inflammation and/or immunological response. In addition, ultrasound can be non-invasively targeted to a specific volume of superficial tissues or deeply embedded organs. Taken together, these properties make sonoporation an innovative and compelling method for gene and drug delivery.




13.5 Principles of Polymerase Chain Reaction (PCR) in cloning and amplifying DNA

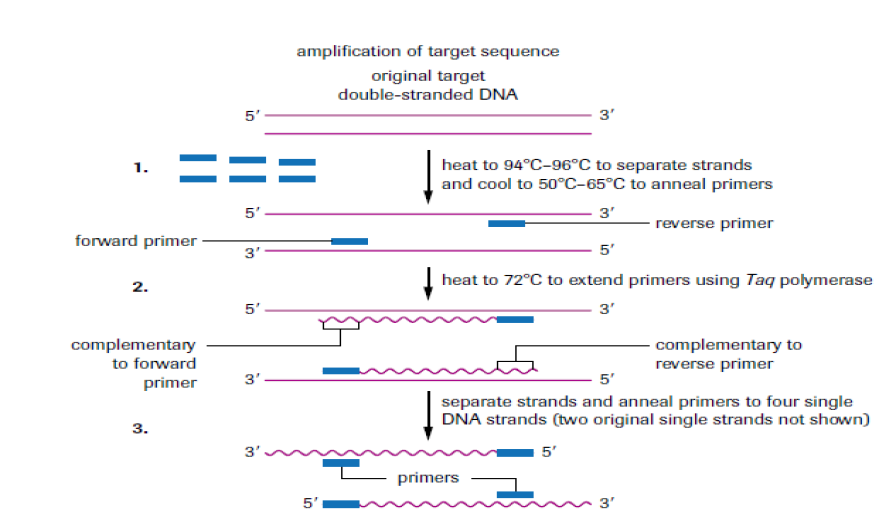
Polymerase chain reaction is a technique that uses the enzyme called DNA polymerase to produce millions of copies of a particular piece of DNA. For simplicity, the two original single strands are not shown after step 3. The main steps of the PCR:
–– The DNA duplex is heated to 90oC to separate the two strands (step 1: denaturation).
–– The mixture is cooled to 60oC to allow the primers to anneal to their complementary sequences (step 2: annealing).
–– At 72oC the primers direct the thermostable DNA polymerase to copy each of the template strands (step 3: extension or elongation of primers).The three steps of the PCR are repeated many times to yield many thousands of copies of the original target sequence. Genes can be cloned by cloning the bacterial cells that contain them, but this requires quite a lot of DNA in the first place. PCR can clone (or amplify) DNA samples as small as a single molecule. It is a newer technique, having been developed in 1983 by Kary Mullis, for his discovery he won the Nobel Prize in 1993.
The polymerase chain reaction is simply DNA replication in a test tube. If a length of DNA is mixed with the four nucleotides (A, T, C and G) and the enzyme DNA polymerase in a test tube, then the DNA will be replicated many times.
Normally, in vivo where DNA replication occurs, the DNA double helix would be separated by the enzymes DNA gyrase and DNA helicase, but in PCR (in vitro) the strands are separated by heating to 95°C for two minutes. This breaks the hydrogen bonds. DNA polymerisation always requires short lengths of DNA (about 20 bases pair long) called primers, to get it started. In vivo the primers are made during replication by DNA polymerase, but in vitro they must be synthesised separately and added at this stage. This means that a short length of the sequence of the DNA must already be known, but it does have the advantage that only the part between the primer sequences is replicated. The DNA must be cooled to 40°C to allow the primers to anneal to their complementary sequences on the separated DNA strands.
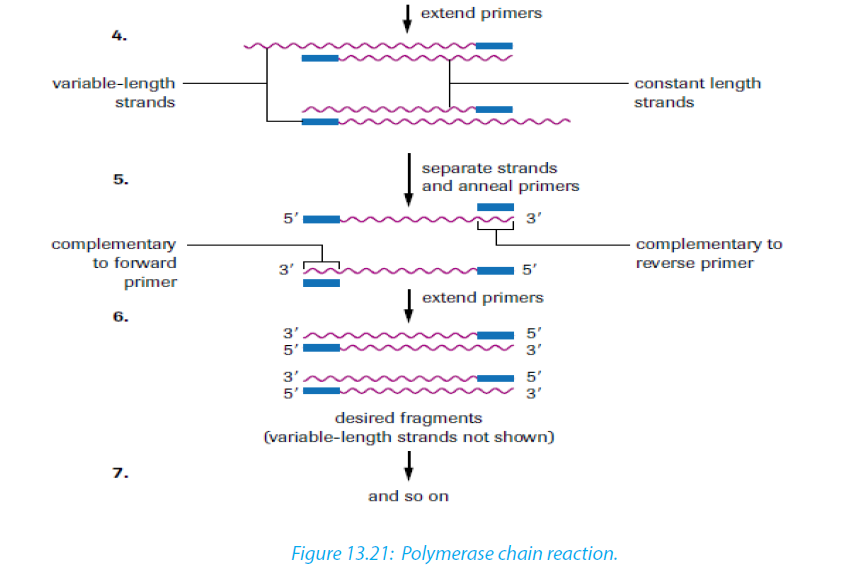
The enzyme (Taq polymerase) used in PCR is derived from the thermophilic bacterium Thermus aquaticus, which grows naturally in hot springs at a temperature of 90°C, so it is not denatured by the high temperatures in step 2. Its optimum temperature is about 72°C, so the mixture is heated to this temperature for a few minutes to allow replication to take place as quickly as possible. Once the primers have annealed, Taq polymerase, a DNA polymerase, can build complementary strands using free nucleotides that have been added to the solution. Each original DNA molecule has now been replicated to form two molecules. The cycle is repeated from step 2 and each time the number of DNA molecules doubles. This is why it is called a chain reaction, since the number of molecules increases exponentially, like an explosive chain reaction. Typically PCR is run from 20 to30 cycles.

Note that:
Artificial DNA synthesis, sometimes known as DNA printing is a method in synthetic biology that is used to create artificial genes in the laboratory. The types of artificial DNA synthesis include recombinant DNA technology, gene purification and PCR (Polymerase Chain Reaction). All of these types have been described above under headings 13.1, 13.3 and 13.5 respectively.

13.6 Gel electrophoresis

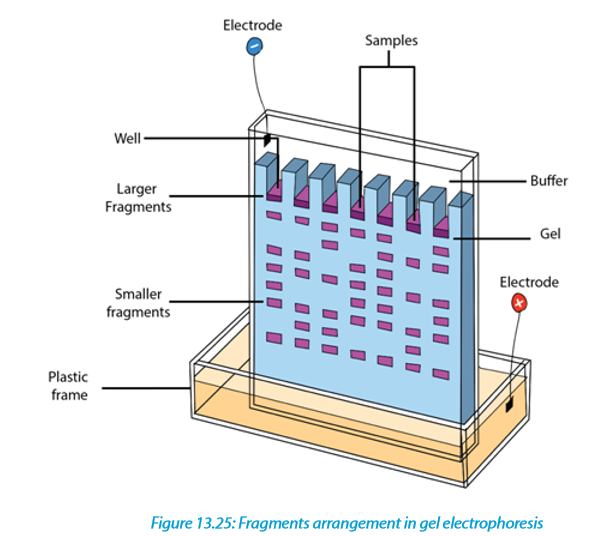
Gel electrophoresis is a laboratory technique used to separate mixtures of DNA, RNA or proteins according to molecular size. In gel electrophoresis, the molecules to be separated are pushed by an electrical field through a gel that contains small pores.
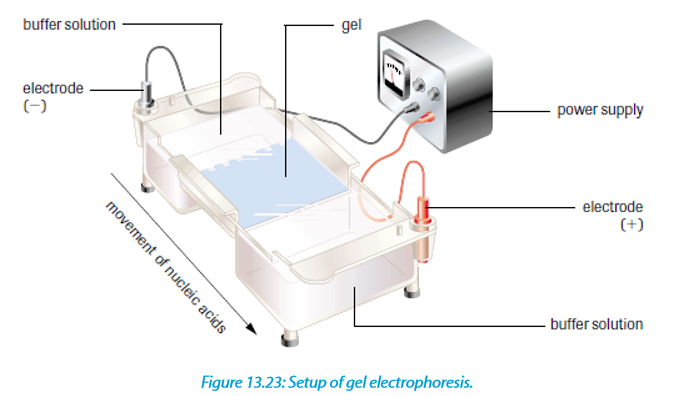
In a common gel electrophoresis setup, a nucleic acid such as DNA is loaded into wells at one end of the gel and then migrates toward the positive electrode at the opposite end. The rate of migration of fragments varies with size. The steps of gel electrophoresis are shown below.
–– The DNA samples are cut with a restriction enzyme into smaller segments of various sizes. The DNA is then placed in wells made on a thick gel.
–– An electric current runs through the gel for a given period of time. Negatively charged DNA fragments migrate toward the positively charged end of the porous gel. Smaller DNA fragments migrate faster and farther than longer fragments, and this separates the fragments by size. The gel floats in a buffer
solution within a chamber between two electrodes.
–– The DNA is transferred to a nylon membrane and radioactive probes are added.
The probes bind to complementary DNA.
–– The X-ray film is exposed to the radiolabelled membrane. The resulting pattern
of bands is called a DNA fingerprint.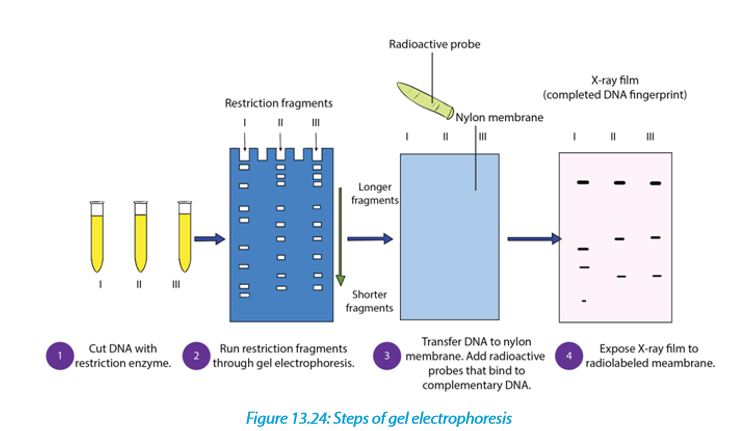
During electrophoresis, DNA fragments migrate through the gel at a rate that is inversely proportional to the logarithm of their size. The shorter the fragment is, the faster it will travel because of its ability to navigate through the pores in the gel more easily than a large fragment can. Larger fragments are hampered by their size. Hence, the longer a nucleotide chain, the longer it takes for the migration.
Gel electrophoresis takes advantage of DNA’s negative charge. Using direct current, a negative charge is placed at one end of the gel where the wells are, and a positive charge is placed at the opposite end of the gel. The electrolyte solution conveys the current through the gel. The negatively charged DNA will migrate toward the positively charged electrode, with the shorter fragments migrating faster than the longer fragments, achieving separation. Small molecules found within the loading dye migrate ahead of all the DNA fragments. Since the small molecules can be visualized, the electrical current can be turned off before they reach the end of the gel.
Once gel electrophoresis is complete, the DNA fragments are made visible by staining the gel. The set of fragments generated with a particular restriction enzyme produces a banding pattern characteristic for that DNA. The most commonly used stain is ethidium bromide. Ethidium bromide is a flat molecule that fluoresces under ultraviolet (UV) light and is able to insert itself among the rungs of the ladder of DNA. When the gel is subjected to UV light, the bands of DNA are visualized because the ethidium bromide is inserted among the nucleotides. The size of the fragments is then determined using a molecular marker as a standard. The molecular marker, which contains fragments of known size, is run under the same conditions (in the same gel) as the digested DNA.
Gel electrophoresis is not limited to the separation of nucleic acids but is also commonly applied to proteins. Proteins are usually run on polyacrylamide gels, which have smaller pores, because proteins are generally smaller in size than nucleic acids. Proteins, however, are not negatively charged; thus, when researchers want to separate proteins using gel electrophoresis, they must first mix the proteins with a detergent called sodium dodecyl sulfate. This treatment makes the proteins unfold into a linear shape and coats them with a negative charge, which allows them to migrate toward the positive end of the gel and be separated. Finally, after the DNA, RNA, or protein molecules have been separated using gel electrophoresis, bands representing molecules of different sizes can be detected. The gel electrophoresis is used for different purposes such as DNA analysis, protein and antibody interactions, testing antibiotics and testing vaccines.

13.7 Use of microarrays in the analysis of genomes and in detecting MRNA

DNA microarray, also commonly known as RNA chip or gene chip or biochip, is technique consisting of a two-dimensional arrangement of DNA molecules representing thousands of cloned genes on a solid surface such as a microscopic slide. DNA microarray shows active genes that are being expression. Since a microarray technology has the potential to examine the expression of several genes at a time, it promises to revolutionize the way scientists study gene expression. For these reasons, DNA microarrays are considered important tools for discovery in clinical medicine.
A basic protocol for a DNA microarray is as follows:
1. Isolate and purify mRNA from samples of interest:
As we are interested in comparing gene expression, one sample usually serves as control, and another sample would be the experiment (for example, a healthy or normal cell versus cancer cell).
2. Reverse transcribe and label the mRNA:
In order to detect the transcripts by hybridization, they need to be labeled, and because starting material may be limited, an amplification step is also used. Labeling usually involves performing a reverse transcription (RT) reaction to produce a complementary DNA strand (cDNA) and incorporating a florescent dye that has been linked to a DNA nucleotide, producing a fluorescent cDNA strand. Disease and healthy samples can be labeled with different dyes and cohybridized onto the same microarray in the following step. Some protocols do not label the cDNA but use a second step of amplification, where the cDNA from RT step serves as a template to produce a labeled cRNA strand.

3. Hybridize the labelled target to the microarray:
This step involves placing labelled cDNAs onto a DNA microarray where it will hybridize to their synthetic complementary DNA probes attached on the microarray. A series of washes are used to remove non-bound sequences. In molecular biology, a hybridization probe is a fragment of DNA or RNA of variable length (usually 100– 1000 bases long) which can be radioactively labelled. It can then be used in DNA or RNA samples to detect the presence of nucleotide sequences (the DNA target) that are complementary to the sequence in the probe.4. Scan the microarray and quantitate the signal:
The fluorescent tags on bound cDNA are excited by a laser and the fluorescently labelled target sequences that bind to a probe generate a signal. The total strength of the signal depends upon the amount of target sample binding to the probes present on that spot. Thus, the amount of target sequence bound to each probe correlates to the expression level of various genes expressed in the sample. The signals are detected, quantified, and used to create a digital image of the array.